
Tous les organismes se composent de cellules. Dans la hiérarchie de l'organisation biologique, la cellule représente le premier niveau capable de vie.
D'ailleurs, bien des être vivants ne sont constitués que d'une seule cellule. Les organismes supérieurs, dont les végétaux et les animaux, comportent plusieurs sortes de cellules spécialisées, organisées en groupements complexes, comme les tissus et les organes.
Cependant, même lorsqu'elles s'unissent à d'autres pour atteindre un niveau d'organisation supérieur, les cellules demeurent toujours les unités fondamentales de la structure et du fonctionnement des organismes. Tout ce que réalise un organisme, il le doit d'abord et avant tout à son activité cellulaire.
Techniques permettant l'étude de la cellule
L'évolution de la science est souvent tributaire de l'invention d'instruments qui permettent à l'être humain d'aller au-delà des limites de ses sens. Ainsi, la découverte et l'étude de la cellule auraient été impossibles sans l'invention et le perfectionnement des microscopes au XVIIe siècle. Encore aujourd'hui, on ne peut étudier la cellule sans utiliser toutes sortes de microscopes.
Microscopie
Les microscopes photoniques (MP) sont appelés ainsi parce que leur source d'éclairage est en général la lumière visible, qui se quantifie en photons. Dans ces instruments, la lumière traverse la préparation (l'échantillon), puis des lentilles de verre. Les lentilles réfractent (dévient) la lumière de façon à grossir l'image projetée dans l'œil. La plupart des structures cellulaires, ou organites, sont invisibles au microscope photonique.
Le microscope électronique, fait passer un faisceau d'électrons à travers la préparation. Les microscopes électroniques modernes atteignent une limite de résolution environ 1000 fois plus grande que celle du microscope photonique. Les biologistes utilisent l'expression ultrastructure cellulaire pour désigner l'anatomie de la cellule que le microscope électronique permet d'observer.
Les deux facteur les plus importants en microscopie sont le grossissement et le pouvoir de résolution. Le grossissement représente le rapport entre les dimensions apparentes de l'image et les dimensions réelles de l'objet. Le pouvoir de résolution est une mesure de la clarté de l'image.
Fractionnement cellulaire
Le fractionnement cellulaire consiste à décomposer les cellules de manière à isoler les principaux organites et à en étudier les fonctions respectives. La centrifugeuse, un instrument capable de tourner à différentes vitesses, sert au fractionnement.
La première étape du fractionnement est l'homogénéisation, qui désintègre les cellules. On cherche généralement à briser les cellules sans trop endommager les organites. La centrifugation de l'homogénat donne un culot (une fraction sédimentée) et un surnageant (une fraction non sédimentée). On recueille le culot pour analyse et on décante le surnageant, après quoi on le centrifuge à nouveau. On recommence le procédé (ultracentrifugation) en augmentant chaque fois la vitesse de centrifugation, et on recueille des constituants cellulaires de plus en plus petits.
Le fractionnement permet d'isoler des constituants cellulaires en grande quantité en vue d'étudier leur composition et leur métabolisme.
Organisation cellulaire
Cellules procaryotes et eucaryotes
Les cellules procaryotes sont exclusives au règne des monères, qui comprend les bactéries et les cyanobactéries. Les cellules eucaryotes, elles, composent les organismes des quatre autres règnes : les protistes, les végétaux, les mycètes et les animaux.
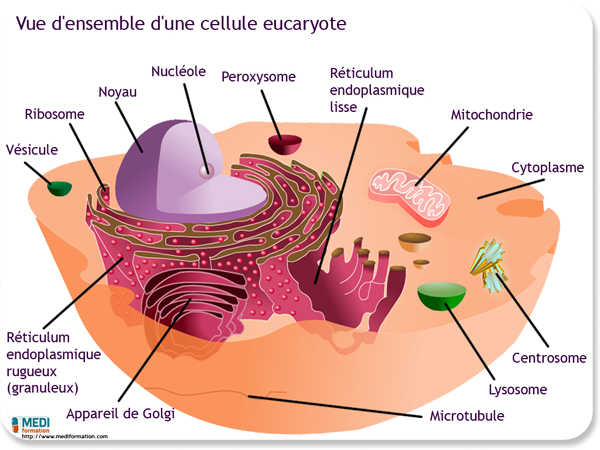
La cellule procaryote n'a pas de noyau véritable. Quand à la cellule eucaryote, elle renferme un noyau véritable contenu dans une enveloppe nucléaire constituée de deux membranes. Toute la région comprise entre cette enveloppe nucléaire et la membrane qui entoure la cellule s'appelle cytoplasme. Celui-ci se compose d'une matière semi-liquide, le cytosol, dans laquelle baignent des organites aux formes et aux fonctions spécialisées. Le cytosol comprend environ 85% d'eau, des glucides, des lipides, des protéines et diverses sortes d'ARN, le tout conférant au cytosol une certaine viscosité. La plupart des organites contenus dans la cellule eucaryote n'existent pas dans la cellule procaryote.
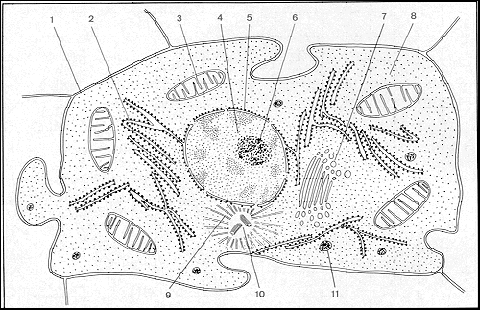
1. membrane 4. nucléoplasme 7. appareil de Golgi 10. centrosomes
2. reticulum endopl. et ribosomes 5. membrane nucléaire 8. cytolasme / hyaloplasme 11. lysosome
3. mitochondrie 6. nucléole 9. aster
Taille des cellules
Pour accomplir ses fonctions métaboliques, la cellule ne doit être ni trop petite ni trop grande, Les plus petites cellules connues appartiennent au règne des bactéries et font partie du genre mycoplasma ; leur diamètre varie entre 0,1 et 1,0 mm. La plupart des bactéries mesurent de 1 à 10 mm de diamètre et sont donc environ dix fois plus grosses que les mycoplasmes. Les cellules eucaryotes, elles, ont typiquement un diamètre dix fois plus grand que les bactéries : 10 à 100 mm.
L'enveloppe extérieure d'une cellule s'appelle membrane plasmique. Cette membrane est un sorte de douanier sélectif qui dirige les échanges de substances chimiques entre la cellule et l'environnement. La membrane laisse passer suffisamment d'oxygène, de nutriments et de déchets pour desservir le volume entier de la cellule. Il y a une limite à la quantité d'une substance donnée qui peut traverser 1 mm 2 de membrane par seconde. Ainsi, plus la surface (mm 2) est grande par rapport au volume, plus les échanges satisfont les besoin cellulaires. Donc, la plupart des cellules sont microscopiques parce qu'il s'agit là de la seule façon de posséder suffisamment de surface par rapport à leur volume pour combler leurs besoins.
Importance de la compartimentation
Malgré leur taille le plus souvent microscopique, les cellules eucaryotes ont des besoins immenses, qui commandent une organisation structurale particulière. Voilà pourquoi elles possèdent des membranes internes qui, à la manière de cloisons, divisent la cellule en compartiments. Ces membranes participent aussi directement au métabolisme cellulaire ; beaucoup d'enzymes se trouvent d'ailleurs enchâssées dans les membranes internes. Étant donné que chaque compartiment cellulaire forme une sorte de micro-environnement, des processus incompatibles peuvent se dérouler simultanément dans la cellule.
Bref, les diverses membranes occupent une place fondamentale dans l'organisation complexe de la cellule. En général, les membranes biologiques se composent d'une double couche de phosphoglycérolipides et d'autres lipides associés à diverses protéines. Par exemple, plusieurs enzymes de la respiration cellulaire sont fixées aux membranes internes des mitochondries. Par conséquent, l'étude des cellules se ramène dans une large mesure à l'étude des membranes et des compartiments fonctionnels (organites) qu'elles délimitent. Les différences entre les cellules végétales et les cellules animales quoique non négligeables, sont moins nombreuses que celles qui séparent les cellules eucaryotes des procaryotes.
Noyau
Le noyau contient la plupart des gènes qui régissent la cellule (les autres se trouvent dans les mitochondries et les chloroplastes). Le noyau constitue généralement l'organite le plus visible d'une cellule eucaryote. Le noyau est entouré de l'enveloppe nucléaire, qui sépare son contenu du cytoplasme.
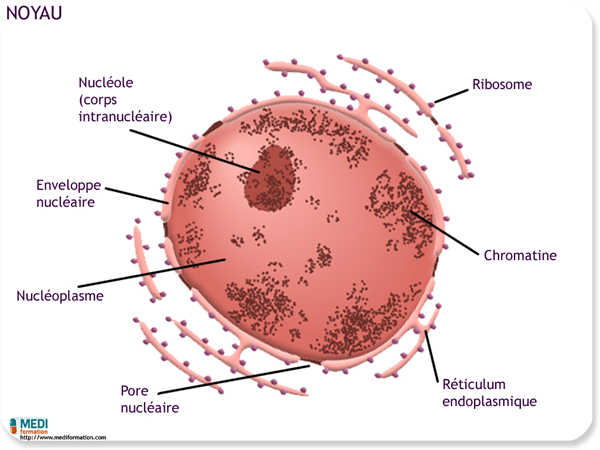
À l'intérieur du noyau se trouve les chromosomes composés d'ADN et de protéines, En dehors des périodes de division cellulaire, les chromosomes sont trop effilés et entremêlés pour qu'on puisse les distinguer individuellement. Au microscope photonique comme au microscope électronique, leur enchevêtrement apparaît comme une masse de matière colorée qu'on appelle chromatine. Les chromosomes deviennent distincts seulement lorsqu'ils se condensent et s'épaississent, au moment où le noyau se prépare à la division.
Entre les périodes de division cellulaire, la structure intranucléaire le plus visible est le nucléole, lieu de synthèse des ribosomes. Le nucléole renferme des segment de chromosomes porteurs de multiples copies des gènes de synthèse des ribosomes, ainsi qu'une quantité considérable d'ARN et de protéines représentant des ribosomes à différentes étapes de leur synthèse ; ces segments du nucléole portent le nom d'organisateurs nucléolaires.
Le noyau régit la synthèse protéique dans le cytoplasme par l'intermédiaire d'un messager moléculaire, l'ARN messager. L'ARN messager (ARNm) est synthétisé dans le noyau selon les directives fournies par l'ADN, puis il sort du noyau par les pores pour apporter les messages génétiques dans le cytoplasme. Là, l'ARNm s'attache à des ribosomes, qui traduisent le message génétique pour élaborer la structure primaire d'une protéine.
Ribosomes
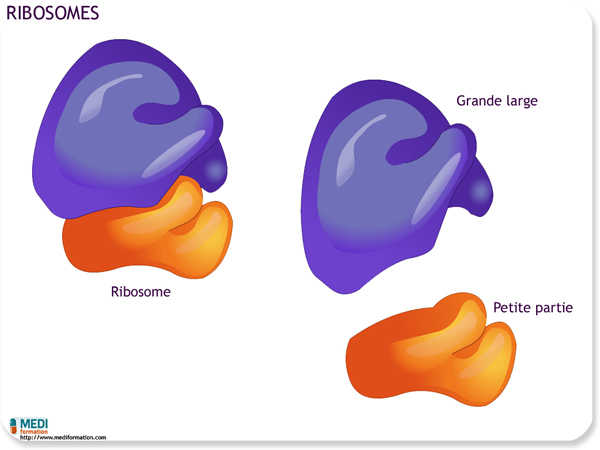
Les ribosomes sont les organites qui assemblent les protéines conformément au code génétique. Ils se comptent par milliers dans une cellule bactérienne et par millions dans une cellule hépatique humaine. Les cellules qui présentent une synthèse protéique intense se démarquent par leur grand nombre de ribosomes, ce qui constitue un autre exemple de la corrélation entre la structure et la fonction.
Il existe deux sortes de ribosomes : les ribosomes libres, en suspension dans le cytosol, et les ribosomes liés, fixés sur le réseau membraneux formant le réticulum endoplasmique. Les ribosomes libres produisent surtout des protéines qui agissent à l'intérieur du cytosol, et ils sont particulièrement abondants dans les cellules en pleine croissance. Les ribosomes liés, quant à eux, synthétisent des protéines destinées à des organites membraneux ou à l'exportation. Les cellules spécialisées dans la sécrétion de protéines, par exemple les cellules du pancréas et d'autres glandes sécrétrices d'enzymes digestives, comportent pour la plupart une forte proportion de ribosomes liés.
Les ribosomes des procaryotes sont plus petits que ceux des eucaryotes, et ils n'ont pas tout à fait la même composition moléculaire.
Réseau intracellulaire de membranes
Les membranes de la cellule eucaryote constituent le réseau intracellulaire de membranes. Ces membranes sont liées de deux façons : ou bien elles sont en continuité les unes avec les autres, ou bien elles s'échangent des portions par l'intermédiaire de vésicules mobiles (sacs membraneux). Toutes les membranes n'ont pas pour autant la même structure et la même fonction. En effet, l'épaisseur, la composition moléculaire et l'activité métabolique d'une membrane peuvent changer à plusieurs reprises au cours de la vie d'une cellule. Le réseau intracellulaire de membranes se compose de l'enveloppe nucléaire, du réticulum endoplasmique, de l'appareil de Golgi, des lysosomes, des peroxysomes, de divers types de vacuoles et de la membrane plasmique (qui n'est pas une membrane interne comme celle des autres organites membraneux, mais qui est tout de même liée au réticulum endoplasmique et aux autres membranes internes).
Réticulum endoplasmique
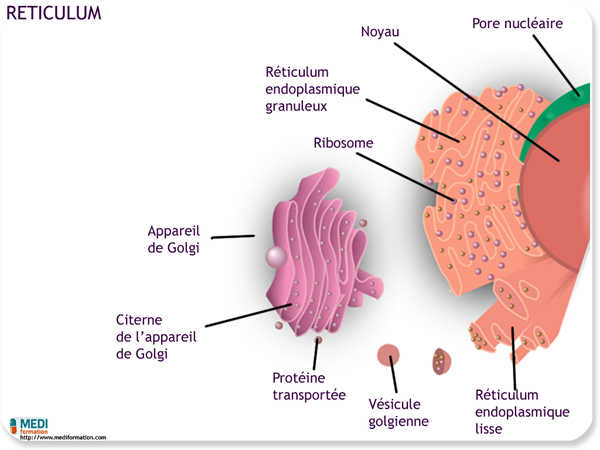
Le réticulum endoplasmique (RE) forme un labyrinthe membraneux si étendu qu'il représente plus de la moitié de toute la substance membraneuse dans beaucoup de cellules eucaryotes. Le réticulum endoplasmique comprend un réseau de tubules et de sacs membraneux appelés citernes. La membrane du réticulum endoplasmique isole du cytosol le contenu des citernes. Et comme elle est en continuité avec l'enveloppe nucléaire, le contenu des citernes communique avec l'espace situé entre les deux membranes de l'enveloppe nucléaire.
Le réticulum endoplasmique se divise en deux régions : le réticulum endoplasmique rugueux et le réticulum endoplasmique lisse. Le réticulum endoplasmique rugueux doit son aspect granulaire aux ribosomes qui parsèment la face cytoplasmique de sa membrane. On trouve aussi des ribosomes sur la face cytoplasmique de la membrane externe de l'enveloppe nucléaire, laquelle s'unit au réticulum endoplasmique rugueux. Le réticulum endoplasmique lisse ne porte pas de ribosomes sur sa face cytoplasmique.
Fonctions du réticulum endoplasmique lisse
Le réticulum endoplasmique lisse participe à divers processus métaboliques, dont la synthèse des lipides, le métabolisme des glucides, ainsi que la détoxication des médicaments, des drogues et des poisons.
Les enzymes du réticulum endoplasmique lisse jouent en effet un rôle important dans la synthèse des graisses, des phosphoglycérolipides, des stéroïdes et d'autres lipides. Parmi les stéroïdes produits par le réticulum endoplasmique lisse, on compte les hormones sexuelles des vertébrés et diverses hormones stéroïdes sécrétées par les glandes surrénales.
Le réticulum endoplasmique lisse joue également un rôle dans le métabolisme des glucides. Les cellules hépatiques nous fournissent un bon exemple de ce rôle. Les cellules hépatique emmagasinent les glucides sous la forme d'un polysaccharide appelé glycogène.
Fonctions du réticulum endoplasmique rugueux
Le réticulum endoplasmique rugueux produit les protéines que beaucoup de cellules spécialisées sécrètent. Les protéines destinées à devenir des produits de sécrétion sont synthétisées par les ribosomes attachés au réticulum endoplasmique rugueux. Lorsqu'un ribosome synthétise une chaîne polypeptidique, celle-ci pénètre la membrane du réticulum endoplasmique, vraisemblablement par un pore. En entrant dans la citerne, la protéine se replie et prend sa configuration native. Puis, avec l'aide d'enzymes enchâssées dans la membrane du réticulum endoplasmique, elle s'unit à un petit polysaccharide par covalence pour devenir une glycoprotéine, comme la plupart des protéines sécrétées.
Une fois les protéines de sécrétion formées, la membrane de réticulum endoplasmique les isole des protéines produites par les ribosomes libres qui, elles, resteront dans le cytosol. Les protéines de sécrétion quittent le réticulum endoplasmique emballées dans des vésicules de transition qui se détachent d'une région spécialisée appelée réticulum endoplasmique de transition.
En plus de participer à la production de protéines de sécrétion, le réticulum endoplasmique rugueux synthétise lui-même ses membranes en jumelant des protéines et des phosphoglycérolipides. Ainsi, grâce à l'agencement de protéines adéquates et de phosphoglycérolipides, le réticulum endoplasmique fait proliférer sa membrane ; ce nouveau matériel peut aussi être transféré, sous la forme de vésicules de transition, à d'autre organites comportant des membranes.
Appareil de Golgi

À leur sortie du réticulum endoplasmique, beaucoup de vésicules de transition se dirigent vers l'appareil de Golgi. On peut comparer l'appareil de Golgi à un centre de fabrication, d'affinage, d'entreposage, de triage et d'expédition. Les produits du réticulum endoplasmique y sont modifiés légèrement et activés, entreposés, puis envoyés vers différentes destinations. Comme vous l'aurez peut-être deviné, l'appareil de Golgi est particulièrement étendu dans les cellules spécialisées dans la sécrétion.
Composé de saccules aplatis, l'appareil de Golgi ressemble à une pile de pains pita. L'appareil de Golgi d'une cellule peut contenir plusieurs empilements reliés en réseau. Chacun des saccules s'entoure d'une membrane qui sépare son contenu du cytosol. Les vésicules de sécrétion, concentrées au voisinage de l'appareil de Golgi, véhiculent des matières entre l'appareil de Golgi et d'autres structures cellulaires.
L'appareil de Golgi présente généralement une nette polarité : les membranes des saccules situés au extrémités opposées d'un empilement n'ont ni la même épaisseur ni la même composition moléculaire. Les deux pôles d'un empilement s'appellent face cis et face trans ; ils ont respectivement pour fonction de recevoir et d'expédier les matières. La face cis est habituellement convexe et située près du réticulum endoplasmique ; elle reçoit ses vésicules de transition. Après s'être détachées du réticulum endoplasmique, les vésicules de transition incorporent leur membrane et leur contenu à la face cis en fusionnant avec la membrane du saccule. La face trans, généralement concave, donne naissance à des vésicules de sécrétion qui s'acheminent vers d'autre sites.
En général, les produits des vésicules de transition du réticulum subissent une modification au cours de leur transit entre la face cis et la face trans de l'appareil de Golgi. Les protéines et les phosphoglycérolipides des membranes formant ces vésicules peuvent également subir une transformation.
L'appareil de Golgi déloge certains monomères des polysaccharides et les remplace par d'autres, produisant ainsi des polysaccharides différents. En plus de ce travail de finition, l'appareil de Golgi fabrique lui-même certaines macromolécules.
Les produits de l'appareil de Golgi destinés à la sécrétion quittent la face trans dans la lumière (cavité) des vésicules de sécrétion, qui fusionneront ultérieurement avec la membrane plasmique.
L'appareil de Golgi élabore et affine ses produits par étapes ; ces étapes correspondent aux différents saccules compris entre la face cis et la face trans, qui renferment chacun des enzymes particulières. Ce sont les vésicules qui s'occupent de faire passer les produits par les saccules successifs.
Lysosomes
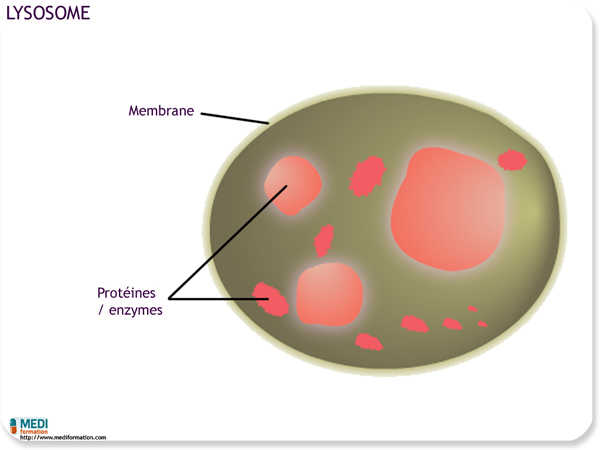
Un lysosome est un sac membraneux rempli d'enzymes hydrolytiques qui digèrent les protéines, les polysaccharides, les lipides et les acides nucléiques ; il s'agit d'un organite animal et non végétal. Le lysosome met la cellule à l'abri des dommages que causeraient les enzymes hydrolytiques si elles circulaient librement dans le cytosol.
Les enzymes hydrolytiques et la membrane du lysosomes sont produites par le réticulum endoplasmique rugueux, puis transférées séparément dans l'appareil de Golgi où leur traitement se poursuit.
La fonction de digestion intracellulaire des lysosomes entre en jeu dans diverses circonstances. Les amibes et beaucoup d'autre protozoaires se nourrissent en englobant de petits organismes et d'autres particules alimentaires, un processus appelé phagocytose. La phagocytose s'observe aussi dans certaines cellules humaines, notamment chez les globules blancs qui englobent les bactéries et les substances étrangères.
Le lysosome a aussi pour fonction de recycler la matière organique intracellulaire, un processus appelé autophagie. Pour effectuer ce recyclage, le lysosome englobe un organite ou un peu de cytosol. Une cellule hépatique humaine, par exemple, recycle la moitié de ses macromolécules chaque semaine. La destruction programmée des cellules par leurs propres lysosomes est une étape importante dans le développement de nombreux organismes.
Vacuoles
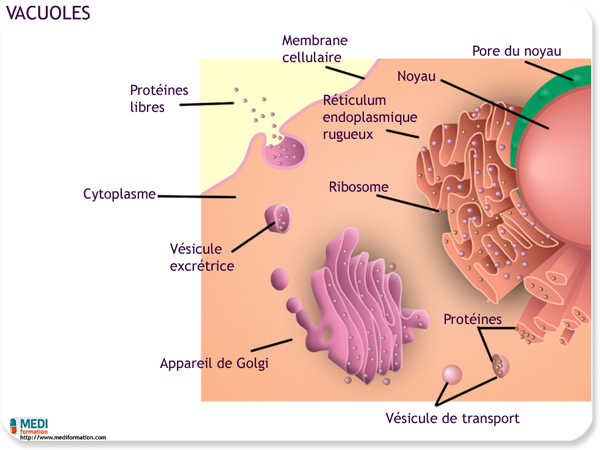
Les vacuoles et les vésicules sont toutes deux des sacs intracellulaires contenus dans une membrane. Plus grosse que les vésicules, les vacuoles ont diverses fonctions. Nous avons déjà traité des vacuoles nutritives, ou phagosomes, formées lors de la phagocytose, et des phagolysosomes formés par la fusion d'un lysosome et d'une vacuole nutritive. Mais les fonctions des vacuoles ne s'arrêtent pas là. Beaucoup de protistes d'eau douce possèdent des vacuoles contractiles qui expulsent l'excès d'eau de la cellule. Les cellules végétales matures contiennent généralement une grande vacuole centrale entourée d'une membrane, le tonoplaste.
La vacuole constitue aussi le principal réservoir d'ions inorganiques tels que les ions potassium et chlorure dans une cellule végétale. Contrairement à la cellule animale, la cellule végétale ne renferme généralement pas de lysosomes spécialisés ; c'est la vacuole qui fait office de compartiment lysosomial. La vacuole sert aussi à l'isolement des sous-produits du métabolisme qui deviendraient nocifs s'ils s'accumulaient dans le cytoplasme. Certaines vacuoles contiennent des pigments, tels les pigments rouges et bleus qui attirent les insectes pollinisateurs vers les pétales des fleurs. Les vacuoles peuvent aussi protéger la plante contre les prédateurs, car elles renferment parfois des composés toxiques ou désagréables au goût. La vacuole joue un rôle primordial dans la croissance de la cellule végétale. En absorbant de l'eau, elle provoque l'allongement de la cellule, qui s'agrandit alors tout en s'épargnant la production de nouveau cytoplasme.
Peroxysomes
Les peroxysomes sont des compartiments métaboliques spécialisés délimités par une membrane simple. Ils contiennent des enzymes qui transfèrent l'hydrogène de divers substrats à l'oxygène. Ils doivent leur nom au sous-produit de ce transfert, le peroxyde d'hydrogène. Les peroxysomes des cellules hépatiques détoxiquent l'alcool et d'autre composés nocifs en transférant l'hydrogène de ces substances à l'oxygène libre.
Dans les graines en germination, les tissus riches en lipides contiennent des peroxysomes spécialisés. Ces organites renferment des enzymes qui déclenchent la conversion des lipides en glucides, un processus qui fournit de l'énergie au jeune plant jusqu'à ce qu'il soit en mesure de produire lui-même ses glucides par photosynthèse.
Les peroxysomes ne proviennent pas des organites membraneux du cytoplasme. Ils se forment en incorporant des protéines et des lipides produits dans le cytosol, et ils se multiplient par scissiparité (division en deux partie égales) quand ils atteignent une certaine taille.
Conversion de l'énergie par les mitochondries et les chloroplastes
Les mitochondries et les chloroplastes sont des organites qui convertissent l'énergie en des formes utilisables par la cellule. Les mitochondries sont le site de la respiration cellulaire, ce processus catabolique complexe qui, à l'aide d'oxygène, produit de l'ATP en extrayant l'énergie des glucides, des lipides et d'autres substances. Les chloroplastes, propres aux végétaux et à certains protistes, convertissent l'énergie solaire en énergie chimique ; avec la lumière qu'ils absorbent, ils alimentent la synthèse de composés organiques à partir de dioxyde de carbone et d'eau. Les mitochondries et les chloroplastes exploitent l'énergie provenant du milieu où vit la cellule.
Les protéines membranaires des mitochondries et des chloroplastes proviennent des ribosomes qu'ils contiennent et des ribosomes libres du cytosol, et non pas du réticulum endoplasmique rugueux. En plus de posséder des ribosomes, les mitochondries et les chloroplastes renferment une petite quantité d'ADN qui programme la synthèse de certaines de leurs protéines. Les mitochondries et les chloroplastes sont des organites semi-autonomes qui croissent et se reproduisent à l'intérieur de la cellule.
Mitochondries
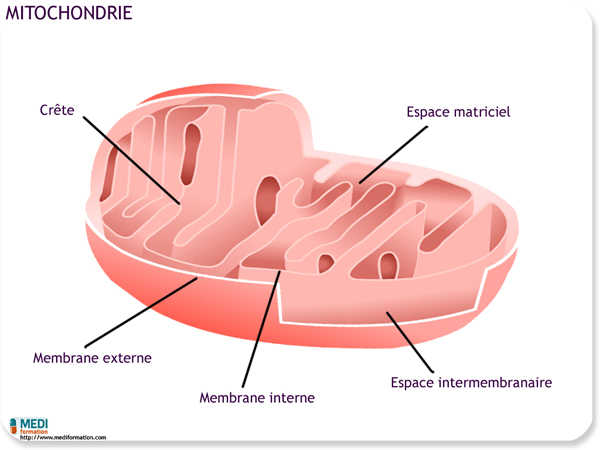
On trouve des mitochondries dans presque toutes les cellules eucaryotes. Certaines cellules ne contiennent qu'une seule grosse mitochondrie, mais la plupart des cellules en comportent des centaines, voire des milliers. Le nombres de mitochondries dépend généralement de l'activité métabolique de la cellule.
L'enveloppe qui entoure la mitochondrie contient deux membranes ; chacune de ces membranes se compose d'une double couche de phosphoglycérolipides dans laquelle s'enchâsse un assemblage unique de protéines. La membrane externe est lisse, mais la membrane interne est repliée sur elle-même et forme des crêtes. Les membranes mitochondriales divisent la mitochondrie en deux compartiments : l'espace intermembranaire, situé entre la membrane interne et la membrane externe, et la matrice mitochondriale, située dans l'espace délimité par la membrane interne. Plusieurs des étapes métaboliques de la respiration cellulaire se déroulent dans la matrice, où diverses enzymes sont concentrées. D'autres protéines nécessaires à la respiration cellulaire, dont l'enzyme qui produit l'ATP, sont intégrées à la membrane interne. Les crêtes de la membrane mitochondriale interne multiplient la surface consacrée à la respiration cellulaire, un autre exemple de corrélation entre la structure et la fonction.
Chloroplastes
Le chloroplaste est un membre spécialisé d'une famille d'organites végétaux appelés plastes. Les amyloplastes (aussi appelés leucoplastes) sont des plastes incolores qui renferment de l'amidon, particulièrement dans les racines et les tubercules. Les chromoplastes, eux, élaborent les pigments qui donnent aux fruit et aux fleurs leur teintes orangées et jaunes. Quant aux chloroplastes, ils contiennent le pigment vert appelé chlorophylle ainsi que d'autres pigments, des enzymes, de l'ADN, de l'ARN, des ribosomes et d'autres molécules nécessaires à la photosynthèse ; de ce fait, les chloroplastes ont une certaine autonomie, comme les mitochondries, et peuvent synthétiser des protéines. Les chloroplastes sont biconvexes et se trouvent dans les feuilles et dans les autres organes verts des végétaux, de même que chez les algues.
Les chloroplastes, les amyloplastes et les chromoplastes naissent des protoplastes présents dans les cellules non spécialisées. Au cours de la croissance de la plante, le sort des protoplastes dépend de la situation des cellules et du milieu dans lequel elles vivent. Dans certaines conditions, les plastes matures changent d'identité.
Le contenu d'un chloroplaste est isolé du cytosol par deux membranes séparées par un espace intermembranaire très mince. À l'intérieur du chloroplaste se trouve un autre réseau membraneux organisé en sacs aplatis, les thylakoïdes. Dans certaines régions du chloroplaste, les thylakoïdes sont empilés comme des jetons de poker et forment des structures appelés grana (granum au singulier). Le liquide où baignent les thylakoïdes s'appelle stroma.
Cytosquelette
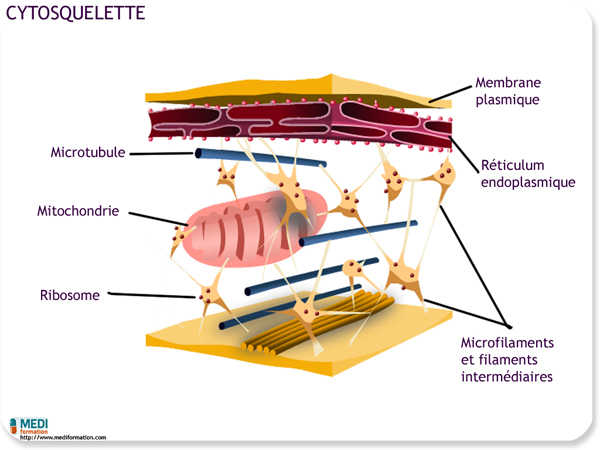
On appelle cytosquelette le réseau de fibres qui parcourt le cytoplasme. Sa fonction consiste à apporter un soutien mécanique à la cellule et à lui conserver sa forme. Cette fonction revêt une importance particulière pour la cellule animale dépourvue de paroi : en s'ancrant au cytosquelette, les organites et certaines enzymes cytoplasmiques restent en place. Le cytosquelette permet à la cellule de changer de forme ; comme un échafaudage, il peut se démonter dans une partie de la cellule et se remonter ailleurs. Le cytosquelette joue aussi un rôle dans la mobilité, qu'il s'agisse du mouvement de la cellule entière ou de celui d'organites à l'intérieur de la cellule. Les fibre du cytosquelette constituent non seulement l'« ossature » de la cellule, mais aussi sa « musculature ». Le cytosquelette sert également à prolonger les pseudopodes (« faux pieds ») de l'amibe et à produire le courant cytoplasmique (la cyclose) dans plusieurs grosses cellules végétales. Le cytosquelette fournit les « monorails » sur lesquels voyagent les vésicules, et ses constituants contractiles manipulent la membrane plasmique pour former les vacuoles nutritives au cours de la phagocytose.
Le cytosquelette comprend au moins trois sortes de fibres. Des plus épaisses aux plus minces, il s'agit des microtubules, des filaments intermédiaires et des microfilaments (aussi appelés filaments d'actine).
Microtubules
Les microtubules sont des tubes rectilignes dont la paroi se compose d'une protéine globulaire, la tubuline.
On trouve des microtubules dans le cytoplasme de toutes les cellules eucaryotes. Dans bien des cas, ils rayonnent d'un centrosome (aussi appelé centre organisateur des microtubules), une masse située près du noyau. Ils servent alors de poutres dans la charpente cellulaire. En outre, des faisceaux de microtubules stratégiquement placés près de la membrane plasmique maintiennent la forme de la cellule. Les microtubules servent aussi de rails sur lesquels les organites peuvent se déplacer et les microtubules participent à la séparation des chromosomes pendant la division cellulaire.
Le centrosome d'une cellule animale contient une paire de centrioles. Chaque centriole comprend neuf triplets de microtubules disposés en cercle. Lorsqu'une cellule se divise, les centrioles se dédoublent. Bien qu'ils concourent probablement à l'assemblage des microtubules, ils ne sont pas essentiels à cette fonction chez tous les eucaryotes. Par exemple, les centrosomes des cellules appartenant au règne des végétaux ne possèdent pas de centrioles.
Cils et flagelles
Les flagelles et les cils, situés à la surface de certaines cellules et servant d'appendices locomoteurs, sont formés par un arrangement spécialisé de microtubules. Beaucoup d'organismes unicellulaires se propulsent dans l'eau au moyen de cils ou de flagelles ; de même, les spermatozoïdes des animaux, ainsi que les gamètes des algues et de certains végétaux sont flagellés. Dans la cellule ciliée ou flagellée qui ne se déplace pas, les cils et les flagelles servent à créer un courant de liquide à la surface du tissu dont fait partie la cellule. Par exemple, les cils des cellules qui tapissent la trachée expulsent des poumons le mucus chargé de débris. Lorsqu'une cellule porte des cils, ceux-ci sont généralement très abondants, alors que les cellules flagellées ne portent habituellement qu'un seul flagelle.
Flagelles et cils ne battent pas de la même façon. Le mouvement d'un flagelle est ondulatoire, et il propulse la cellule dans l'axe du flagelle. Le mouvement ciliaire, en revanche, ressemble à celui d'un avion : il fait alterner un battement de propulsion et un battement de récupération ; l'ensemble des battements de propulsion poussent la cellule perpendiculairement à l'axe du cil.
Microfilaments et mouvement
Les microfilaments sont de forme cylindrique. Ils se composent de molécules d'actine, une protéine globulaire, qui s'unissent pour former des chaînes.
Les microfilaments sont surtout connus pour leur rôle dans la contraction musculaire. Des milliers de microfilaments d'actine sont disposés en parallèle le long de la cellule musculaire, en alternance avec des filaments plus épais composés d'une protéine appelée myosine. La contraction de la cellule résulte du glissement des microfilaments d'actine contre les filaments de myosine.
Aussi concentrés et ordonnés qu'ils soient dans les cellules musculaires, les microfilaments semblent néanmoins présents dans toutes les cellules eucaryotes pour y remplir une fonction de soutien.
Les agrégats d'actine et de myosine sont à l'origine des contractions cellulaires localisées. Chez certaines cellules mobiles, les microfilaments interviennent dans l'élongation et la rétraction des pseudopodes qui permettent à la cellule entière de se déplacer sur une surface. Dans les cellules végétales, les microfilaments concourent à la cyclose, un phénomène par lequel tout le cytoplasme circule sans cesse dans l'espace séparant la vacuole et la membrane plasmique.
Filaments intermédiaires
Les filaments intermédiaires doivent leur nom à leur diamètre qui est supérieur à celui des microtubules. Les filaments intermédiaires comprennent divers éléments cytosquelettiques, dont la composition protéique varie d'un type de cellule à l'autre. À l'opposé, les microtubules et les filament ont le même diamètre et la même composition dans toute les cellules eucaryotes. Les filaments intermédiaires sont aussi plus stables que les microfilaments et les microtubules. Il se pourrait que les divers types de filaments intermédiaires aptes à supporter la tension, constituent l'armature du cytosquelette entier.
Paroi cellulaire
La paroi cellulaire fait partie des caractéristiques distinctives de la cellule végétale. Elle protège la cellule, maintient sa forme et prévient une absorption excessive d'eau. À l'échelle de la plante, la paroi résistante des cellules s'oppose à la gravitation et garde l'organisme à la verticale.
Mesurant de 0,1 à plusieurs micromètres, la paroi cellulaire végétale est beaucoup plus épaisse que la membrane plasmique. La paroi cellulaire présente une composition assez uniforme : elle est constituée de fibres de cellulose enchâssées dans une matrice amorphe faite d'autres polysaccharides et de protéines.
La cellule végétale immature commence par sécréter une paroi relativement mince et flexible, la paroi primaire, à l'aide des microtubules et des vésicules de sécrétion de l'appareil de Golgi. Entre les parois primaires de cellules adjacentes, on trouve la lamelle moyenne, une mince couche riche en polysaccharides adhésifs appelés pectine. La lamelle moyenne, fabriquée par la cellule mère à la fin de la division cellulaire, colle les cellules filles les unes aux autres. Arrivée à maturité, la cellule durcit sa paroi. Pour ce faire, certaines cellules sécrètent simplement des substances raffermissantes dans la paroi primaire. D'autres ajoutent une paroi secondaire entre la membrane plasmique et la paroi primaire. La paroi secondaire, souvent élaborée par apposition de couches successives, a une matrice résistante et durable qui fournit protection et soutien à la cellule.
Glycocalyx des cellules animales
Les cellules animales sont dépourvues de paroi, mais beaucoup d'entre elles montrent en surface une couche duveteuse et légèrement adhésive, le glycocalyx. Le glycocalyx comprend tous les glucides rattachés par liaisons covalentes tant aux protéines qu'aux lipides de la membrane plasmique ; à ces glucides s'ajoutent des glycoprotéines sécrétées par la cellule et qui demeurent près de la surface cellulaire. Le glycocalyx favorise l'adhérence entre les cellules. Certain de ses constituants jouent le rôle d'« étiquettes » qui procurent une identité aux cellules, par exemple aux globules rouges. Certains constituants du glycocalyx font office de récepteurs membranaires. Souvent, l'adhérence que le glycocalyx confère aux cellules animales est augmentée par les jonctions cellulaires situées entre les membranes des cellules adjacentes.
Jonctions intercellulaires
Il arrive souvent que les cellules adjacentes adhèrent, interagissent et communiquent directement par l'intermédiaire de canaux traversant les parois cellulaires, les plasmodesmes. Ces derniers relient les membranes plasmiques et les contenus cytoplasmiques de cellules adjacentes. On trouve trois principaux types de jonction intercellulaire : les jonctions serrées, les desmosomes et les jonctions ouvertes.
EXTRAIT DE : BIOLOGIE , CAMPBELL. ED. DE BOECK