
La vie sur Terre existe grâce à l'énergie solaire. Les chloroplastes des végétaux captent l'énergie lumineuse qui a parcouru les 160 millions de kilomètres environ qui nous séparent du Soleil. Ensuite, ils la convertissent en énergie chimique et ils l'emmagasinent dans des glucides et d'autres molécules organiques, qu'ils forment à partir de dioxyde de carbone et d'eau. Ce processus s'appelle photosynthèse.
La photosynthèse nourrit presque tous les êtres vivants, directement ou indirectement. Un organisme se procure les composés organiques nécessaires à la production d'ATP et de chaînes carbonées par autotrophie ou par hétérotrophie. De prime abord, le terme autotrophe semble contredire le principe suivant lequel les cellules constituent des systèmes ouverts qui tirent leurs ressources de leur milieu. Toutefois, les autotrophes ne sont autosuffisants que dans la mesure où ils ne mangent ni ne décomposent d'autres organismes. Ils élaborent leurs molécules organiques à partir de matières premières inorganiques. Les végétaux sont autotrophes.
Plus précisément, les végétaux sont photoautotrophes, c'est-à-dire qu'ils utilisent la lumière comme source d'énergie pour synthétiser des glucides, des lipides et des protéines. La photosynthèse s'observe aussi chez les algues et certains autres protistes et chez quelques procaryotes.
Incapables de produire eux-mêmes leur nourriture, les hétérotrophes se nourrissent de composés synthétisés par d'autres organismes. Presque tous les hétérotrophes, l'être humain y compris, ont absolument besoin des photoautotrophes, non seulement pour se nourrir mais également pour respirer, car l'oxygène est un sous-produit de la photosynthèse. Par conséquent, la nourriture que nous ingérons et l'oxygène que nous respirons proviennent du chloroplaste.
Le chloroplaste : site de la photosynthèse
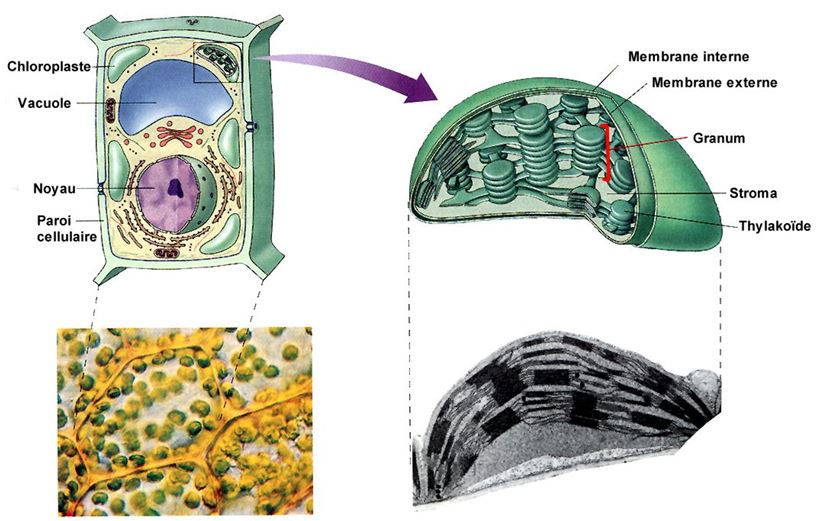
Toutes les parties vertes d'une plante, y compris les tiges vertes et les fruits non encore mûrs, comprennent des chloroplastes, mais ce sont généralement les feuilles qui en renferment le plus. La couleur de la feuille vient de la chlorophylle, le pigment vert contenu dans les chloroplastes. Les chloroplastes abondent tout particulièrement dans le mésophylle, le tissu interne de la feuille. Le dioxyde de carbone entre dans la feuille et l'oxygène en sort par des pores microscopiques appelés stomates. L'eau absorbée par les racines, elle, se rend dans les feuilles par les nervures. Les nervures servent également à transporter les glucides jusqu'aux parties non photosynthétiques de la plante.
L'enveloppe extérieure du chloroplaste se compose de deux membranes. À l'intérieur du chloroplaste se trouve un liquide dense, le stroma, où baignent des sacs membraneux aplatis appelés thylakoïdes. La membrane des thylakoïdes délimite un compartiment appelé espace intrathylakoïdien . Ici et là, les thylakoïdes forment des empilements denses appelés grana (granum au singulier). La chlorophylle se trouve dans les membranes des thylakoïdes.
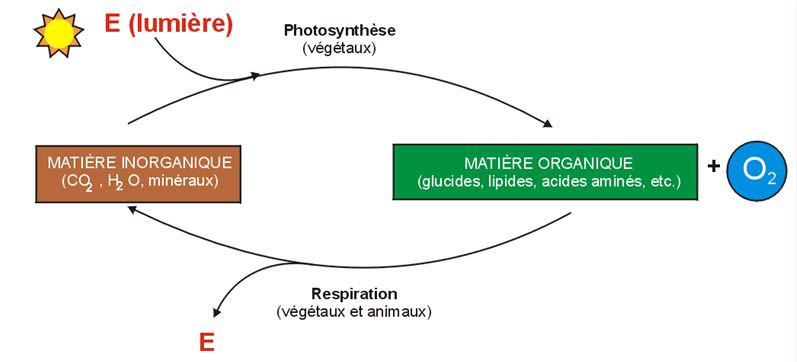
Aperçu de la photosynthèse
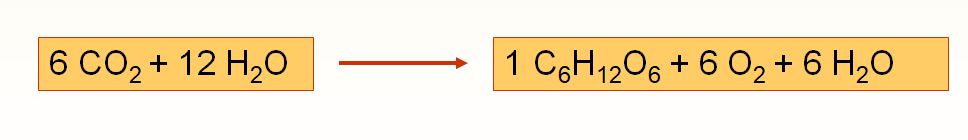
Cette équation chimique simplifiée révèle que le changement chimique réalisé pendant la photosynthèse est l'inverse de celui qui a lieu pendant la respiration cellulaire. La cellule végétale est le siège de ces deux processus métaboliques.
Scission de la molécule d'eau
Le principal résultat du brassage d'atomes réalisé pendant la photosynthèse est l'extraction de l'hydrogène de l'eau et son incorporation au glucide.
Le résidu de la photosynthèse, l'oxygène, «remplace» l'oxygène atmosphérique consommé pendant la respiration cellulaire.
Les deux phases de la photosynthèse
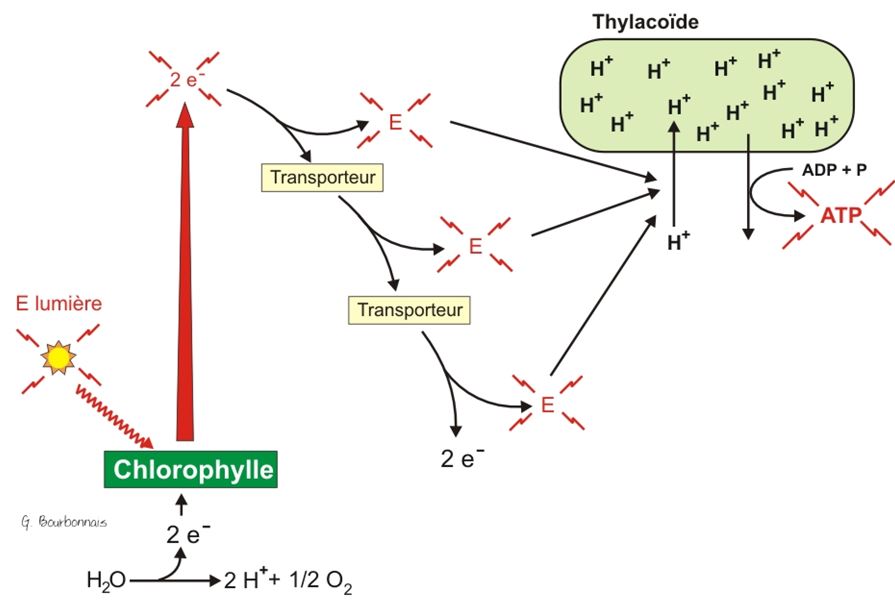
L'équation de la photosynthèse, sous des apparences de simplicité, présente un processus fort complexe. De fait, la photosynthèse comprend deux phases, elles-mêmes divisées en de nombreuses étapes. Ces deux phases sont les réactions photochimiques et le cycle de Calvin, aussi nommé phase de fixation du carbone. Les réactions photochimiques se déroulent dans les thylakoïdes du chloroplaste, tandis que le cycle de Calvin a lieu dans le stroma.
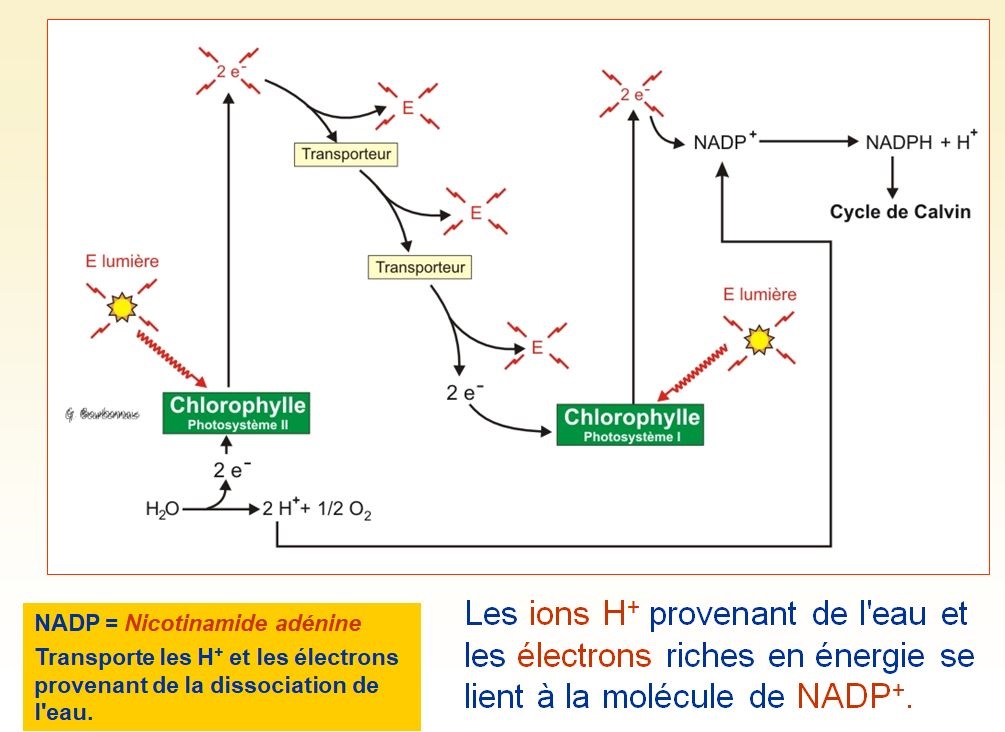
Les réactions photochimiques incluent les étapes de la photosynthèse qui convertissent l'énergie solaire en énergie chimique. La lumière absorbée par la chlorophylle déclenche un transfert d'électrons et de protons de l'eau vers un accepteur appelé NADP+ (nicotinamide adénine dinucléotide phosphate), qui stocke temporairement les électrons riches en énergie. La molécule d'eau se trouve ainsi scindée ; par conséquent, ce sont les réactions photochimiques qui rejettent de l'oxygène.
Les réactions photochimiques utilisent l'énergie solaire pour réduire le NADP+ en NADPH + H+ en lui ajoutant une paire d'électrons et deux protons (H+). De plus, les réactions photochimiques produisent de l'ATP, car elles alimentent l'ajout d'un groupement phosphate à l'ADP, un processus appelé photophosphorylation. Par conséquent, la conversion initiale de l'énergie lumineuse en énergie chimique donne deux composés : le NADPH + H+, une source d'électrons riches en énergie, et l'ATP, la devise énergétique des cellules.
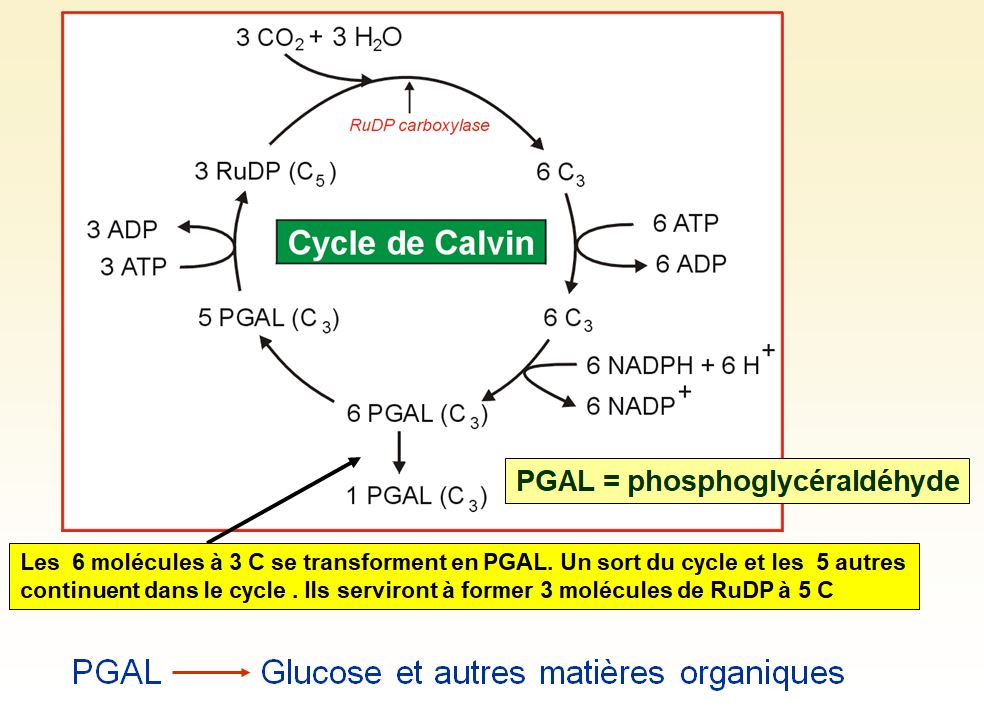
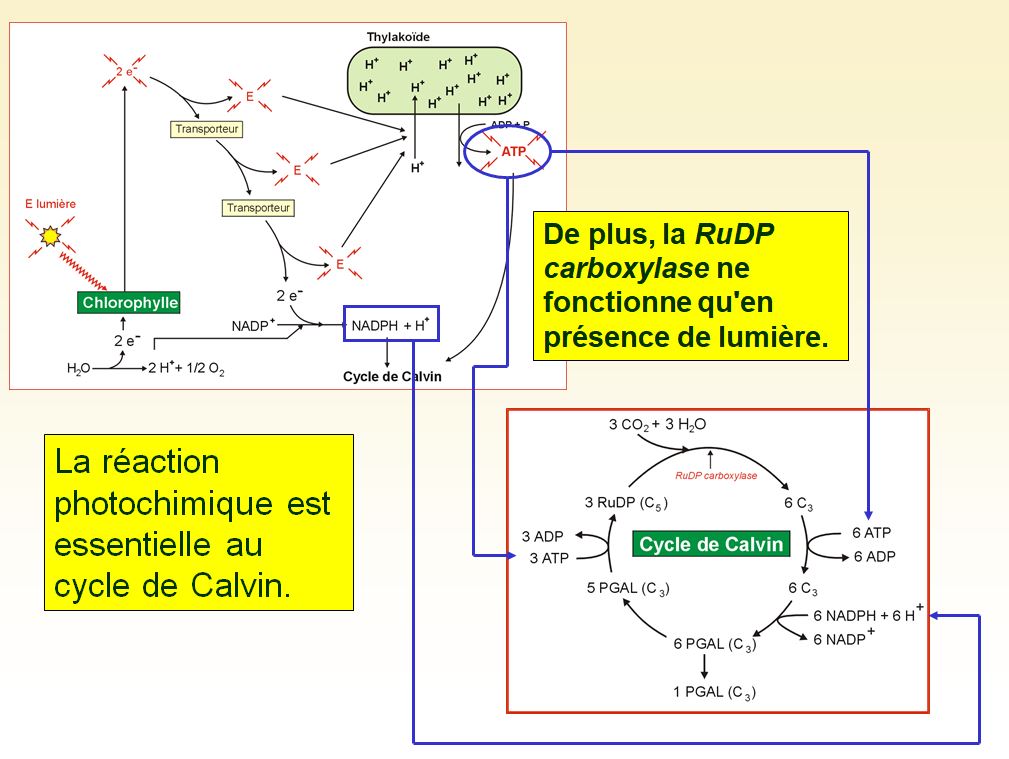
Le cycle de Calvin commence par l'incorporation de dioxyde de carbone atmosphérique aux molécules organiques déjà présentes dans le chloroplaste. On appelle cette étape fixation du carbone. Le carbone fixé se fait ensuite réduire en glucide par l'ajout d'électrons et de protons. Le potentiel réducteur provient du NADPH + H+, qui a acquis des électrons riches en énergie pendant les réactions photochimiques. Pour convertir le dioxyde de carbone en glucide, le cycle de Calvin a aussi besoin d'énergie chimique sous forme d'ATP, également produite pendant les réactions photochimiques.
Le carbone entre dans le cycle de Calvin sous la forme de dioxyde de carbone, et il en sort sous forme de glucide. Le glucide produit n'est pas du glucose mais un glucide à trois atomes de carbone appelé phosphoglycéraldéhyde (PGAL). Pour synthétiser une mole de ce glucide, le cycle doit fixer trois moles de dioxyde de carbone et, partant, se dérouler trois fois.
Le cycle de Calvin attache chaque mole de dioxyde de carbone à une mole d'un glucide à cinq atomes de carbone appelé ribulose diphosphate (RuDP en abrégé). L'enzyme qui catalyse cette première étape est la RuDP carboxylase, la protéine la plus abondante dans les chloroplastes et probablement sur terre. La réaction donne un intermédiaire à six atome de carbone, si instable qu'il se scinde aussitôt en deux moles de 3-phosphoglycérate. À l'étape suivante, chaque molécule de 3-phosphoglycérate reçoit un groupement phosphate supplémentaire pris à l'ATP par une enzyme, ce qui forme du 1,3-phosphoglycérate en PGAL. Plus précisément, les électrons du NADPH + H+ réduisent le groupement carboxyle du 1,3-phosphoglycérate, qui devient alors le groupement carbonyle du PGAL ; ce PGAL est un glucide à trois carbones, le même que celui que la glycolyse forme en scindant le glucose.
Le cycle a commencé avec un capital de glucide valant 15 moles de carbone, c'est-à-dire avec trois moles de ribulose diphosphate à cinq atomes de carbone. Maintenant, on compte 18 moles de carbone sous la forme de six moles de PGAL. Cependant, une seule de ces moles de PGAL compte pour un gain net en glucide. En effet, une mole sort du cycle pour être utilisée par la cellule végétale, mais les cinq autres doivent aller régénérer les trois moles de ribulose diphosphate. Au cours d'une série complexe de réactions, les dernières étapes du cycle réarrangent les chaînes de carbone des cinq moles de PGAL en trois moles de ribulose diphosphate. Pour ce faire, le cycle dépense trois autres moles d'ATP. Le ribulose diphosphate est alors de nouveau prêt à recevoir du dioxyde de carbone. Le cycle recommence.
Pour synthétiser une mole nette de PGAL, le cycle de Calvin consomme neuf moles d'ATP et six moles de NADPH + H+. Les réactions photochimiques régénèrent l'ATP et le NADPH + H+. Le PGAL issu du cycle de Calvin devient la matière première de voies métaboliques qui synthétisent d'autres composés organiques, dont une variété de glucides.
Les étapes métaboliques du cycle de Calvin sont parfois appelées réactions obscures, car aucune ne nécessite directement de la lumière. Chez la plupart des végétaux, néanmoins, le cycle de Calvin se déroule pendant le jour, car il s'agit du seul moment où les réactions photochimiques peuvent régénérer le NADPH + H+ et l'ATP utilisés par la réduction du dioxyde de carbone en glucide. Essentiellement, le chloroplaste produit des glucides à l'aide de l'énergie lumineuse en coordonnant les deux phases de la photosynthèse.
Pigments photochimiques
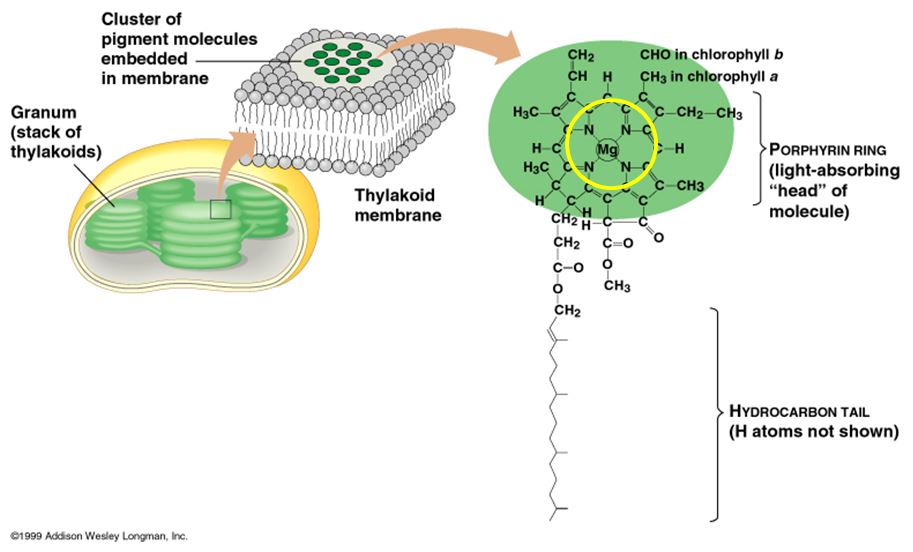
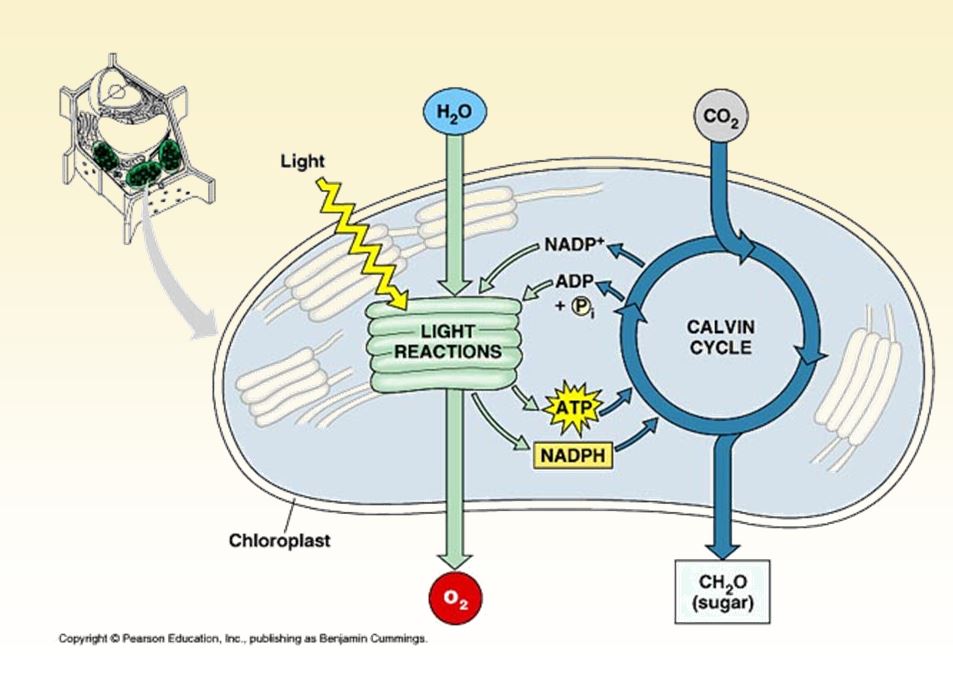
Lorsque la lumière rencontre la matière, celle-ci peut la réfléchir, la transmettre ou l'absorber. Les substances qui absorbent la lumière visible s'appellent pigments. Chaque pigment absorbe une gamme de longueurs d'onde différente et la fait ainsi disparaître. Les feuilles nous paraissent vertes parce que la chlorophylle absorbe la lumière rouge et la lumière bleue en même temps qu'elle réfléchit et transmet la lumière verte. La lumière bleue et la lumière rouge sont donc les plus favorables à la photosynthèse, tandis que la lumière verte est la moins propice.
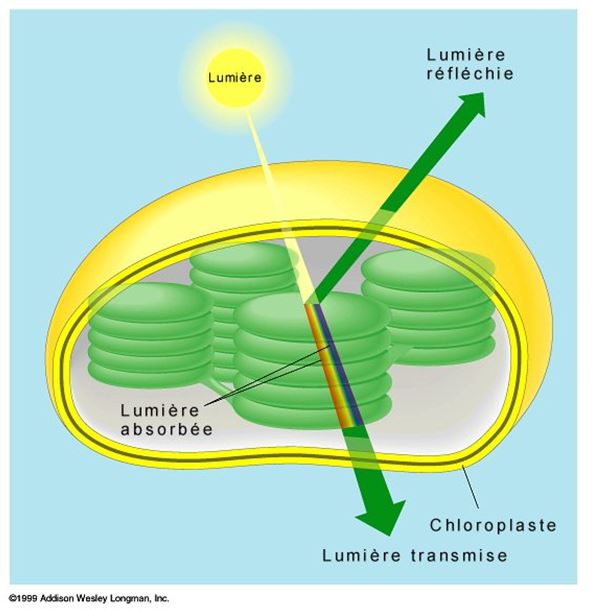
La chlorophylle a et la chlorophylle b sont presque identiques, mais la légère différence structurale qui les distingue suffit à leur donner des spectres d'absorption différents, et par le fait même, des couleurs distinctes. La chlorophylle a est bleu-vert, tandis que la chlorophylle b est jaune-vert. Le chloroplaste renferme aussi une famille de pigments accessoires appelés caroténoïdes (carotène), dont la couleur varie du jaune à l'orangé.
Photosystèmes (en lecture)
Dans la membrane des thylakoïdes, la chlorophylle a, la chlorophylle b et les caroténoïdes se trouvent arrangés en différents amas de plusieurs centaines de molécules. Parmi les nombreuses molécules de chlorophylle a de chaque amas, seule une paire précise de molécules peut déclencher les réactions photochimiques en cédant ses électrons excités à l'accepteur primaire. L'emplacement de ces molécules spécialisées de chlorophylle a dans les amas de pigments se nomme centre réactionnel. Les autres molécules de chlorophylle a ainsi que les molécule de chlorophylle b et de caroténoïdes forment une antenne de captation qui absorbe les photons et transmet l'énergie de la molécule jusqu'au centre réactionnel. Tout l'appareil, constitué par l'antenne, la chlorophylle du centre réactionnel et l'accepteur primaire d'électrons, s'appelle photosystème. Les photosystèmes sont les unités photoréceptrices de la membrane des thylakoïdes.
La membrane des thylakoïdes comprend deux types de photosystèmes : le photosystème I et le photosystème II. Le centre réactionnel du photosystème I se compose d'une paire de molécules de chlorophylle a appelée P700 ; ce pigment doit son appellation au fait qu'il absorbe la lumière ayant une longueur d'onde de 700 nm, située dans la partie rouge du spectre. Le centre réactionnel du photosystème II, lui, consiste en une paire de molécules de chlorophylle a appelée P680 ; le spectre d'absorption de ce pigment culmine à 680 nm. En fait, les pigments P700 et P680 sont des molécules de chlorophylle a identiques mais associées à des protéines différentes ; la distribution de leurs électrons et leurs spectres d'absorption diffèrent donc légèrement. Le P700 et le P680 ne se démarquent que par leur position dans la membrane des thylakoïdes ; ils sont liés à des protéines particulières et se trouvent très proches de leurs accepteurs primaires respectifs.
Transport cyclique d'électrons
Au cours des réactions photochimiques, le transport d'électrons peut se faire selon deux trajets : cyclique ou non cyclique. Le transport cyclique d'électrons est le trajet le plus simple. Il ne fait intervenir que le photosystème I et n'engendre que de l'ATP ; il ne produit ni NADPH + H+ ni oxygène. Cette voie est dite cyclique parce que les électrons excités qui quittent la chlorophylle au centre réactionnel finissent par y revenir. A chaque réaction d'oxydoréduction survenant dans la chaîne de transport, les électrons perdent de l'énergie potentielle, pour trouver finalement leur état fondamental dans le P700.
A mesure que les électrons excités perdent de l'énergie en retournant au P700 via la chaîne de transport, la membrane des thylakoïdes exploite cette perte énergétique des électrons pour actionner la synthèse de l'ATP. La chaîne de transport d'électrons achemine des protons à travers la membrane des thylakoïdes, ce qui crée une force protonmotrice. Une ATP synthétase analogue à l'ATP synthétase mitochondriale se sert de cette force protonmotrice pour produire de l'ATP. On emploie le terme photophosphorylation pour désigner la synthèse d'ATP dans les chloroplastes, car elle est amorcée par l'énergie lumineuse. La production d'ATP au cours de transport cyclique d'électrons s'appelle photophosphorylation cyclique.
Transport non cyclique d'électrons
Le transport non cyclique d'électrons, au cours duquel les électrons passent continuellement de l'eau au NADP+, fait intervenir les deux photosystèmes. Comme dans le transport cyclique, la lumière excite les électrons du P700, le centre réactionnel du photosystème I. Cependant, les électrons ne retournent pas au centre réactionnel : ils sont mis en réserve dans le NADPH + H+. Le NADPH + H+ jouera ultérieurement le rôle de donneur d'électrons et de protons, quand le cycle de Calvin réduira le dioxyde de carbone en glucide.
La chlorophylle oxydée devient elle-même un agent oxydant très puissant ; les « trous » laissés par ses électrons doivent être comblés. C'est ici que le photosystème II entre en jeu : il remplace les électrons du centre réactionnel du photosystème I. Quand l'antenne du photosystème II absorbe la lumière, l'énergie atteint le P680, la chlorophylle a spécialisée du centre réactionnel. L'accepteur primaire d'électrons du photosystème II piège les électrons éjectés du P680 et les transfère à une chaîne de transport, celle-là même qui participe au transport cyclique d'électrons. Les électrons dévalent la chaîne, perdant de l'énergie potentielle en cours de route, jusqu'à ce qu'ils atteignent le P700 et remplissent les vides laissés quand le photosystème I a réduit le NADP+. À mesure que les électrons dérivent du photosystème II au photosystème I, la chaîne de transport achemine des protons à travers la membrane des thylakoïdes. La force protonmotrice peut alors actionner la synthèse de l'ATP. La production d'ATP au cours du transport non cyclique d'électrons est appelée photophosphorylation non cyclique. Soulignons cependant que la synthèse de l'ATP se fait de la même façon que dans la photophosphorylation cyclique.
Jusqu'ici, le transport non cyclique d'électrons a produit du NADPH + H+ et de l'ATP et il a remplacé les électrons perdus dans le centre réactionnel du photosystème I. Or, le P680, la chlorophylle du centre réactionnel du photosystème II, a maintenant des vides à combler, et les électrons de substitution proviendront de l'eau. Une enzyme extrait des électrons de l'eau et les fournit au P680. Le retrait des électrons scinde la molécule d'eau en deux protons et un atome d'oxygène, lequel se combine immédiatement avec un autre atome d'oxygène pour former de l'oxygène moléculaire (O2).
Résumons maintenant les réactions photochimiques. Le transport non cyclique d'électrons pousse les électrons de l'eau, où ils possèdent peu d'énergie potentielle, vers le NADPH + H+, où ils renferment beaucoup d'énergie potentielle. Le courant d'électrons que la lumière engendre produit en outre de l'ATP. Par conséquent, l'équipement de la membrane des thylakoïdes convertit l'énergie lumineuse en énergie chimique emmagasinée dans le NADPH + H+ et dans l'ATP. L'oxygène constitue un sous-produit des réactions photochimiques.
À quoi sert la photophosphorylation cyclique ? Nous constatons que le transport cyclique constitue un court-circuit ; quand les électrons éjectés du P700 atteignent la ferrédoxine, il se font renvoyer en direction de la molécule de chlorophylle plutôt que de se faire aiguiller vers le NADP+. Le transport non cyclique d'électrons produit des quantités à peu près égales d'ATP et de NADPH + H+, mais le cycle de Calvin consomme plus d'ATP que de NADPH + H+. La photophosphorylation cyclique comble la différence en produisant de l'ATP mais pas de NADPH + H+.
Facteurs externes influant sur la photosynthèse
En milieu naturel, certains facteurs environnementaux influent sur l'intensité de la photosynthèse.
La concentration molaire volumique du CO2 dans l'air ne limite pas la photosynthèse, malgré la tendance actuelle à l'effet de serre. In vitro, toutefois, une augmentation graduelle du CO2, jusqu'à cinq fois la teneur normale, entraîne un accroissement graduel de la photosynthèse. Par ailleurs, la disponibilité de l'eau intervient régulièrement et à toute latitude.
La lumière, pour sa part, exerce son influence de diverses manières. L'intensité de la photosynthèse augmente proportionnellement à l'intensité de l'éclairement, puis atteint un maximum qui varie selon les espèces. In vitro, un éclairement ininterrompu de plusieurs jours ralentit la photosynthèse, car les produits de synthèse viennent à engorger les chloroplastes. En milieu naturel, la composition de la lumière blanche reste constante jour après jour, et ce facteur n'influe pas comme tel sur l'activité photosynthétique durant l'année. In vitro, cependant, une plante verte réagit davantage à certaines longueurs d'onde dont celles correspondant à la lumière rouge et à la lumière bleue. Elle fournit un meilleur rendement avec la lumière blanche qui regroupe l'ensemble des longueurs d'onde du spectre visible.
Il existe un lien étroit entre la lumière et la température ambiante. Plus cette dernière augmente, plus elle favorise la photosynthèse, jusqu'à la température extrême du fonctionnement des enzymes. La valeur nutritive du milieu exerce également une influence ; s'il s'appauvrit jusqu'à ralentir la croissance de la plante ou à modifier la structure des chloroplastes, la photosynthèse décroît d'autant.
Extrait de CAMPBELL, DeBoeck Université
Autres sites: